Video sobre la importancia del fuego en los brezales andaluces, esos matorrales mediterráneos dominados por brezo (Calluna vulgaris) en suelos pobres en nutrientes. Gran parte de las especies de plantas de estos brezales necesitan incendios para reproducirse.
Realizado por Guillermo Ojeda, con la colaboración de Fernando Ojeda, Susana Gómez, Universidad de Cádiz, Plan INFOCA, Parque Natural de los Alcornocales, etc.
Prescribed burns has only been introduced in the Valencia region (eastern Iberia) very recently (end of 2019), and are undertaken by the Valencia government. One of the first burns was performed Castell de Castells (Alacant province) in March 2021, it was of relatively low intensity (compare with natural wildfires in the area). The area was dominated by a mediterranean shrublands with few young pines (P. halepensis). One year latter the area is a paradise of flowers; below are a few of them. Thank you to J. Fabado and X. Riera (Jardí Botànic de Valencia) for their help in the species names.
Castell de Castells burn, March 2021
Castell de Castells, May 2022
First row: Reseda (alba) valentina, Tulipa australis, Sarcocapnos enneaphylla / Second row: Linaria_depauperata, Teucrium homotrichum_ronniger, Neotinea_maculata / Third row: Argyrolobium zanonii, Phlomis lychnitis, Teucrium pseudochamaepitys / Fourth row: Anagallis arvensis, Asphodelus cerasiferous, Aphyllantes monspeliensis. Photos by JG Pausas except Linaria by B. Moreira.
El pasado 24 de abril falleció un amigo, el Profesor Welington Delitti, después de luchar duramente, y durante más de dos años, contra un cáncer. Welington era profesor de ecología en el Instituto de Biociencias de la Universidad de Sao Paulo (Brasil), donde dirigió a numerosos estudiantes; también tuvo un papel importante en la dirección del Instituto (director 2007-2011 y subdirector 2011-2015). Su investigación se centraba principalmente en el cerrado (sabanas) de Brasil, con especial énfasis en los ciclos de materia orgánica y nutrientes y, más recientemente, en la restauración y conservación de ese maravilloso ecosistema. Durante su visita a Valencia (España) trabajó sobre el efecto de la recurrencia del fuego en matorrales de coscoja (Quercus coccifera). Hizo una aportación importante a la ecología del cerrado, y muchos de sus trabajos fueron publicados en revistas científicas brasileñas e internacionales (ver lista de publicaciones aquí). Y fue él quien me mostró ese ecosistema tan interesante por primera vez.
Personalmente era una persona entrañable, buena, y muy comprometida con la universidad y la sociedad. Era muy apreciado en su universidad por su buena gestión del Instituto de Biociencias. Su carácter afable y tranquilo le llevaba a resolver los conflictos de la forma más socialmente correcta posible. Nos enseñó una manera de funcionar, y por ello, su legado es grande.
Welington Delitti en un Cerrado de Sao Paulo, Brasil 2008 (foto: JG Pausas)
Fotos de Welington Delitti tomadas entre 2008 y 2021. La foto de arriba-derecha, por Marcos Santos (USP Imagens, 2013); las 3 fotos de la fila de abajo y la del medio-izquierda, extraídas del Facebook de Welington (2018-2012); las otras 3 fotos son propias (2008, 2014; JG Pausas).
Ecology and evolutionary biology have focused on how organisms fit the environment. Less attention has been given to the idea that organisms can also modify their environment, and that these modifications can feed back to the organism, thus, providing a key factor for their persistence and evolution [1]. We propose that there are at least three independent lines of evidence emphasising these biological feedback processes at different scales (figure below): niche construction (population scale); alternative biome states (community scale); and the Gaia hypothesis (planetary scale). Flammability is an example of niche construction [2], and the forest-savanna mosaics are an example of the alternative biome states [3] (figure below).
The importance of feedback processes make us rethink traditional concepts like niche and adaptation. For instance, the idea of evolution as a process of adaptation to fit a pre-existing environment needs to be replaced by a ‘co-evolutionary’ species-environment approach. An implication is that the concept of species niche, and niche occupancy, is less relevant than traditionally thought. That is, organisms do not adapt to a pre-existing environment (available niche), they construct their environment and then both ‘co-evolve’. A higher level of fitness is the result of this coevolution. Feedbacks also provide an alternative framework for understanding spatial and temporal patterns of vegetation that differ from those based on gradual changes (e.g., gradient analysis and succession), and suggest that multi-stability and abrupt transitions in a given environment are common [3]; this also has implications for species’ niche modelling [4].
Earth is in transition to a new and warmer state due to anthropogenic forcing, and feedback thinking may help us understand the process. We suggest that incorporating feedback thinking and understanding how feedbacks may operate at different scales may help in opening our minds to key processes contributing to the dynamics and resilience of our biosphere.
Fig. 1. Examples of eco-evolutionary feedbacks at different organising levels: Niche construction (population; e.g. flammability), alternative biome states (community; forests and savannas) and Gaia (biosphere). The signs of the feedbacks indicate the most common type of feedback for each example. Evolutionary feedbacks represent the evolutionary processes at the different scales (from selection at the micro-evolutionary scale to the acquisition of key macro-evolutionary innovations). From [1].
References
[1] Pausas J.G. & Bond W.J. 2022. Feedbacks in ecology and evolution. Trends Ecol. Evol. [doi | pdf]
[2] Pausas J.G., Keeley J.E., Schwilk D.W. 2017. Flammability as an ecological and evolutionary driver. J. Ecol. 105: 289-297. [doi | wiley | pdf]
[3] Pausas J.G. & Bond W.J. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
[4] Pausas J.G. & Bond W.J. 2021. Alternative biome states challenge the modelling of species’ niche shifts under climate change. J. Ecol. 109: 3962-3971. [doi | wiley | pdf]
Many plants concentrate their seedling recruitment after the passage of a fire. This is because postfire conditions are especially optimal for germination and establishment of many species as fires create extensive vegetation gaps that have high resource availability, minimal competition, and low pathogen load. Thus we propose that fireprone ecosystems create ideal conditions for the selection of seed dormancy as fire provides a mechanism for dormancy release and optimal conditions for germination [1]. We compiled data from a wide range of fire-related germination experiments for species in different ecosystems across the globe and identified four dormancy syndromes: heat-released (physical) dormancy, smoke-released (physiological) dormancy, non-fire-released dormancy, and non-dormancy. In fireprone ecosystems, fire, in the form of heat and/or chemical by-products (collectively termed ‘smoke’), are the predominant stimuli for dormancy release and subsequent germination, with climate (cold or warm stratification) and light sometimes playing important secondary roles. Fire (heat or smoke)-released dormancy is best expressed where woody vegetation is dense and fires are intense, i.e. in crown-fire ecosystems (e.g., mediterranean-type ecosystems). In grassy fireprone ecosystems (e.g. savannas), where fires are less intense but more frequent, seed dormancy is less common and dormancy release is often not directly related to fire (non-fire-released dormancy). Fire-released dormancy is rare to absent in arid ecosystems and rainforests. Heat-released dormancy can be traced back to fireprone floras in the ‘fiery’ mid-Cretaceous, followed by smoke-released dormancy, with loss of fire-related dormancy among recent events associated with the advent of open savannas and non-fireprone habitats. Anthropogenic influences are now modifying dormancy-release mechanisms, usually decreasing the role of fire. We conclude that contrasting fire regimes are a key driver of the evolution and maintenance of diverse seed dormancy types in many of the world’s natural ecosystems.
Fig. 1. Percentage germination of 68 populations or species subjected to simulated fire- (y axis) and summer-type (warm stratification) temperature (x-axis) (C., Cistus; F., Fumana; U., Ulex; A., Acacia; M., Mimosa). Points above the dotted line (1:1) have higher germination levels after fire heat than after summer heat. Note that all points at or below the line are for species in savannas [S], while the others are from mediterranean shrublands and other crown-fire ecosystems. That is, in crown-fire ecosystems, fire is the most likely selective agent for dormancy. From [1].
Fig. 2. Dated phylogeny for major clades in the New and Old World Cistaceae together with closely related ancestral clades. Pie charts at the tips show the fraction of species that occur in crown-fire ecosystems (red), surface-fire ecosystems (orange), those with physical dormancy – hard seeds (green), and those with heat-released dormancy (blue). Blank sectors mean that the trait is absent. Letters at the tips refer to growth forms in the clade (T, tree; S, shrub or subshrub; H, herb/annual). Black dots indicate the crown age of diversification of the corresponding clade. From [1].
References
[1] Pausas J.G. & Lamont B.B. 2022. Fire-released seed dormancy – a global synthesis. Biological Reviews [doi | pdf | supp. mat. | data (figshare)]
El pasado sábado 5 de marzo falleció Antoni Escarré Esteve a los 80 años y en su tierra natal, Alacant. Era Doctor en Biología por la Universitat de Barcelona, catedrático de ecología en la Universitat d’Alacant, y fundador del departamento de ecología de esa universidad. Escarré fue uno de los pioneros de la ecología terrestre en nuestro país, amigo muy cercano a los ecólogos catalanes más eminentes (Ramón Margalef, Jaume Terradas). Su pasión naturalista le llevó a trabajar con una gran diversidad de organismos tanto animales como plantas. Realizó trabajos clave para entender el ciclo de la materia y nutrientes, la ecofisiología de plantas, y la fauna en ecosistemas mediterráneos; posteriormente se enfocó en ecosistemas tropicales. Nunca dejó de mostrar curiosidad por la naturaleza, incluso en su jubilación, ya alejado de obligaciones docentes o investigadoras. Su última tesis doctoral se leyó en 2019.
Una de sus actividades destacadas fue la de dirigir un doctorado sobre desarrollo sostenible del bosque tropical en Cuba, en el cual se han realizado un centenar de tesis doctorales. Este programa ha sido fundamental para el desarrollo de la ecología en Cuba. Por ello Escarré es muy conocido y apreciado por todo naturalista cubano, y fue investido Doctor Honoris Causa por la Universidad de Pinar del Río en Cuba en 2007.
Antoni Escarré también participó en la vida política de la Comunidad Valenciana y ostentó diversos cargos políticos durante los periodos socialistas, tales como Conseller d’Educació, Cultura i Ciència (1989-1991) y posteriormente, Conseller de Medi Ambient (1991-1993). Se dice que era una persona demasiado sincera y normal (es decir, informal para los políticos) para la política, y pronto volvió a la docencia en la UA. Entre sus aficiones destacaba el hockey, fue jugador y entrenador, y llegó a ser Presidente de la Federación de Hockey de la Comunidad Valenciana (1986).
A nivel personal era encantador, culto, bueno, sincero, alegre, comprometido, generoso e inteligente; Terradas, con quien compartió numerosas vivencias, decía que era imposible aburrirse con él. En los últimos años lo hemos visitamos varias veces en su casa, y siempre estaba jovial, buscando cualquier excusa para celebrar algo e invitarnos a una copa de cava. Las veladas en su casa, donde discutíamos de ciencia y política, a menudo en compañía de alguien de Cuba, eran entrañables.
Le echaremos de menos
La primera fotografía (en blanco y negro): A. Escarré cuando era conseller de cultura (1990), en compañía de Enric Valor. El resto de fotografías: Antoni Escarré con algunos de sus colaboradores del doctorado cubano: Vicente Berovides, Rubén Chamizo y Josabel Belliure
No soy experto en conflictos, ni en Rusia, ni en Ucrania, pero no puedo evitar expresar mi opinión. Las reacciones y razonamientos que se dan por muchos políticos y medios de comunicación me parecen extremadamente pobres.
1) Condeno rotundamente la invasión de Ucrania por parte de Rusia. El gobierno ruso se comporta de manera extremadamente imperialista, autócrata, e inhumana (igual que lo hizo en Siria, e igual que EEUU en Irak, y en tantos otros sitios).
2) Estoy en contra de que se solucionen conflictos y desacuerdos mediante violencia, y especialmente mediante guerras. Los líderes que proponen guerras son los menos perjudicados, los ciudadanos de a pie, los más perjudicados. No es cuestión de pacifismo, es cuestión de que las guerras enquistan los conflictos, no los solucionan.
3) Ante una acción violenta, hay que apoyar a los oprimidos, independientemente de quien tenga más razón en el conflicto. El desequilibrio de fuerzas es desproporcionado, es un abuso de superioridad.
4) Solidaridad con los ciudadanos en Rusia que están manifestandose en contra de esta guerra; arriesgan mucho. Y solidaridad con los desertores del ejercito ruso; arriesgan muchísimo. Hay que evitar que aparezca una rusofobia; estamos en contra de las decisiones del gobierno ruso, no de los ciudadanos. Tanto en Rusia como en Ucrania (y España) hay fascistas, comunistas, demócratas, neoliberales, anarquistas, ecologistas, apolíticos, y un largo etc.
5) Esta guerra es un fracaso de la humanidad, y en especial de Europa y EEUU, ya que no se ha conseguido (¿o querido?) evitar, y hasta el momento no se ha sabido detener. Había margen de maniobra antes de que empezara. Los tímidos intentos por algunos políticos (p.e., Macron, etc.) no fueron suficientes, y dadas las posibles consecuencias, se debería haber hecho mucho mucho más esfuerzo en negociar una solución. Quizá podría haber ayudado contar con un panel de expertos en conflictos, con premios Nobel de la Paz, con la ONU, o con reconocidos líderes mundiales (Papa, Dalai Lama), etc. Justificar la guerra diciendo que con Putin no se puede negociar es de una pésima calidad democrática. Es conocido desde hace mucho tiempo que Rusia no quiere bases de la OTAN en su frontera, igual que EEUU no quieres pro-rusos en la suya (Cuba). ¿Qué haría EEUU si México quisiera instalar una base militar rusa? ¿Es necesaria la OTAN? Es difícil imaginar que haya valido la pena empeñarse en que la OTAN tenga bases militares en Ucrania. ¿Dónde está la ONU?
6) Estoy en contra de enviar armamento a Ucrania, que sólo alimenta la guerra y no soluciona el conflicto. La cantidad de armamento que requeriría Ucrania para “ganar” a Rusia es ingente. Además, enviando armas hay peligro de que la guerra se “mundialice” e incluso de que se use armamento nuclear. Todos los esfuerzos se deben centrar en parar la guerra, no en alimentarla. Enviar armas es una acción fácil y una manera de lavarse las manos; pensar que con eso se está ayudando al pueblo Ucraniano es engañoso y peligroso. La guerra no la ganará ni Putin ni Ucrania, la perderemos todos. Además, se alimenta el comercio y la corrupción que hay en torno al negocio del armamento y las guerras. ¿Enviamos armas a Palestina?
7) Los que dicen que la izquierda apoya a Rusia desconocen lo que significa ser de izquierdas en el siglo XXI.
8) Celebro que Europa facilite la acogida de refugiados (pero demuestra que si no se realizó lo mismo en otros conflictos previos fue por falta de ética o directamente por racismo).
9) Tengo mis dudas de que las sanciones impuestas por la Unión Europea a Rusia sean realmente eficaces porque: (1) afectarán a la población rusa (no sólo a los responsables de la guerra); (2) la Unión Europea tiene una gran dependencia comercial de Rusia (gas, cereales, minería, etc.), y esta podría tomar represalias contra Europa (guerra económica); y (3) Rusia es una gran potencia militar y está geográficamente muy cerca (en la misma Europa), por lo que no parece muy inteligente enfrentarse a ella.
10) A menudo los medios de comunicación cuentan la guerra como si fuera una película, presentando a los ucranianos casi como héroes que están defendiendo valientemente sus causas y resistiendo los ataques. La verdad es que cada día hay más destrucción y muertos en Ucrania, y más desplazados, mientras el ejército ruso sigue avanzando inexorablemente.
La pérdida de suelo es de las peores cosas que le puede suceder a un ecosistema, ya que tarda muchísimo en recuperarse. Por ello, después de un incendio, en los sitios donde hay posibilidad de pérdida de suelo, se aconseja realizar medidas rápidas de protección del suelo, tales como poner paja, ramas, haces de ramas, ramas trituradas, troncos en fajinas, o cordones de restos de cortas. Estas acciones reducen el impacto de las gotas de agua de la lluvia, generan condiciones de humedad para la regeneración de la vegetación, y reduce el movimiento del suelo. En una zona incendiada, la extensión de zonas con potencial de erosión depende, especialmente, del tipo suelo, la pendiente, y el uso previo de la zona. En las zonas sobre calizas duras de la costa mediterránea, las extensiones con potencial de erosión posincendio suelen ser reducidas, a veces puntuales; nunca en todo un incendio.
Actualmente estamos viendo que después de incendios, se están realizando fajinas con troncos y ramas en zonas donde no se espera pérdida de suelo, tales como zonas planas, pedregosas, incluso en bancales con muros bien conservados (ver fotos abajo). Para ello se cortan los pinos y algunos arbustos (a veces incluso especies rebrotadora) y se amontonan a modo de fajinas. Estas acciones parecen un gasto económico poco justificable, además de perder los beneficios de los árboles quemados en pie.
Cabe recordar que los árboles muertos en pie benefician a la regeneración porque también disminuyen el impacto de las gotas de lluvia en el suelo, mantienen cierta humedad, captan agua de la niebla, y sirven de posadero para aves que traen semillas y que contribuyen a la regeneración. Además de ser hábitat para mucha fauna (principalmente invertebrados y algunas aves). Cuando los árboles muertos caen, proporcionan materia orgánica y nutrientes al suelo. Sería necesaria una justificación para cortarlos, y para concentrar la biomasa en pocos puntos (ver fotos).
Las siguientes fotos corresponden a ejemplos de fajinas de troncos y ramas realizadas después de incendio, en zonas que no se espera erosión por estar en bancales, en zonas pedregosas, o en zonas con poco pendientes. Fotos tomadas en dos incendios de la Comunidad Valenciana: Llutxent (noviembre de 2018) y Azuébar (diciembre de 2021).
This post complements the letter published today in the Science [doi] – A version of that letter is also available in Turkish here.
Turkey was hit hard by wildfires in 2021, with a record of about 203,000 ha burnt. Most of the area burnt was covered by Mediterranean Pinus brutia forests. Pinus brutia is not a fire-resistant trees, it dies after a fire; however, they have serotinous cones thus after fire the seeds are dispersed and new individuals recruit few months later. This forest also includes many shrubs able to resprout or germinate after fire. Thus natural regeneration was expected in most of the affected area. In fact, 4 months after fire, we already observed pine seedlings and many species resprouting [link]. To preserve this ecosystems, it is important to preserve their regeneration potential. Usually, quick postfire management is only needed if soil losses are likely; in those environments, soil losses typically occurs in only a small proportion of the landscape.
However, the Turkish government is cutting all dead trees (salvage logging). In many places, heavy machinery is being used and forest roads are being opened. In some cases, logging is followed by seeding or by terracing and new tree planting. That is, in some places they are transforming an ecosystem to an artificial afforestation. Thus the postfire management actions are more disturbing than the fire. And these postfire actions are taking place in both unprotected public forests and in conservation areas (e.g. Marmaris National Park).
It is worth remembering that standing dead trees have many ecological functions such as to reduce the impact of raindrops on the ground (i.e., reducing erosion), maintain some humidity, capture water from fogs, serve as perches for birds that bring seeds and contribute to the regeneration, and are habitat for many fauna (mainly invertebrates and some birds). And when dead trees fall down, they provide organic matter and nutrients to the soil.
We urge the Turkish General Directorate of Forestry to stop degrading ecosystems and move toward more ecologically sustainable forest management.
All photos below were Pinus brutia forests.
Postfire salvage logging + terracing + plantation in Marmaris National Park (see also this video)
Postfire salvage logging of burned trees in the Marmaris area
Examples of destroying potential natural postfire regeneration in Antayla, Turkey. Click the image to enlarge. Photos: link
References
[1] Tavsanoglu, Ç. & Pausas J.G: 2022. Turkish postfire action overlooks biodiversity. Science [doi | pdf | Turkish version]
The role of large herbivores in explaining broad-scale ecological pattern has often been underestimated [1]. Plants have defenses against large herbivores (e.g., spines, high wood density [2]). And many continents had abundant large herbivores (megafauna) that were extinguished in Pleistocene (except in Africa). In a recent paper [3] we asked, to what extent the past distribution of extinct magafauna explains current geographical distribution of plant defense traits in the Neotropics (South & Central America). We fond that a significant proportion of the variance in the distribution of wood density, leaf size, stem spines, and leaf spines are explained by variable related to past megafauna (richness and body mass).
We defined 3 antiherbiomes in South America, that is, regions with characteristic plant defenses, environmental conditions, and Pleistocene megafauna, as follows: Small-Leaves-Thorny (SLT): thorny and small-leaved plants, in arid, cold and nutrient-rich ecosystems, containing numerous extinct and extant large grazers. Intermediate-Leaves-Woody (ILW): intermediate leaf sizes and levels of chemical defenses, and very high wood density, in moist and hot climates, and extremely nutrient-poor soils; and a high extinct megafauna richness, especially in relation to small browsers and mixed-feeders. Broad-Chemically-defended-Leaves (BCL): very large leaves with chemical defenses, mostly associated with moist climates and intermediate fertility soils, with few but large extinct megafauna species, especially browsers. Similar antiherbiomes can be observed in current Africa. These antiherbiomes represent one of the most striking broad-scale anachronisms in ecology.
We estimated that in South America, savannas occupied about 10 millions of Km2 during the Pleistocene, ca. 63% of them were converted to forests (44% to moist forests, 19% to dry forests) after the megafauna extinction (biome shifts [4]), and ca. 37% remains as savanna (stable). This suggests that South America was a savanna-dominated continent, much more similar to Africa than today, and that a large proportion of South American forests are the result of megafauna extinctions.
Overall our results suggest that past (extinct) large herbivores explain an important proportion of the variability of current plant traits and community assemblies.
Fig. 1. Left: Distribution of the 3 anti-herbiomes. Right: Hypothesized distribution of savanna during the Pleistocene (coloured areas; based on the distribution of extinct megafauna), that currently are savanna (in yellow), moist forests (dark green) and dry forests (light green). From [3]
Fig. 2. Reconstruction of Pleistocene savanna (ILW antiherbiome) with Taxodon platensis (a mixed feeder) next to the tree Bowdichia virgilioides (sucupira-preto; Fabaceae), and a Notiomastodon in the background. Artist: Júlia d’Oliveira
Fig. 3. Additional reconstructions of the Pleistocene Brazilian savannas from [5]. Artist: Júlia d’Oliveira
References
[1] Pausas JG & Bond WJ. 2019. Humboldt and the reinvention of nature. J. Ecol. 107: 1031-1037. [doi | jecol blog | jgp blog | pdf]
[2] Dantas V & Pausas JG. 2020. Megafauna biogeography explains plant functional trait variability in the tropics. Glob. Ecol. & Biogeogr. [doi | pdf | data:dryad | blog ]
[3] Dantas V., Pausas J.G. 2022. The legacy of Southern American extinct megafauna on plants and biomes. Nature Comm. 13: 129 [doi | pdf | data & codes] – New!
[4] [2] Pausas JG & Bond WJ. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
[5] Pansani et al. 2019. Isotopic paleoecology (δ13C, δ18O) of Late Quaternary megafauna from Mato Grosso do Sul and Bahia States, Brazil. Quat Sci Rev, 221, 105864.
During last summer, over 170,000 ha burnt in Turkey. One of these fires was the Marmaris fire, a fire of about 12,500 ha in SW of the country. The area includes part of the Marmaris National Park and an Special Environmental Protection Area. Most of the area were covered by Pinus brutia forests, a Mediterranean pine that has some serotinous cones.
Four months after the Marmaris fire, I visited the area together with Çagatay Tavsanoglu. Some plants were resprouting and some geophytes flowering (Cyclamen, Arisarum). Pine seedlings had started to germinate; there were also many other seedlings but still too small to identify them (e.g., Cistus species). Below are a few examples of plants that were regenerating after the fire (click on the photos to enlarge them).
El pasado día 10 de noviembre impartí una conferencia titulada ‘Fuego: Incendios y biodiversidad’ en la delegación del CSIC de Valencia, con motivo de la celebración de los 25 años del CIDE (Centro de Investigaciones sobre Desertificación). Aquí la tenéis disponible.
hora:minuto
0:00 a 0:04 – Presentación por el director del CIDE 0:04 a 0:47 – Conferencia propiamente dicha 0:47 a 1:24 – Preguntas del público
It is very nice to see that COP26 have decided to halt deforestation and accelerate the restoration of natural ecosystems (link). We hope this time it will be effective and quick. Past attempts such as REDD+ or the 2014 New York Declaration on Forest (PDF) had limited success; deforestation (logging and clearing for farming or mining) continued even in regions with high biodiversity value (e.g., the Amazon rainforests, the savannas of Cerrado, the dry forest of the Chaco, and Indonesian closed-canopy tropical forests; Figs. 1,2,3,4 below). Deforestation and forest fragmentation are also the main reasons behind the large number of fires occurring in rainforests (native rainforests are non-flammable).
We also hope the COP26 decision will stimulate the conservation not only of forests but also other ecosystems with high biodiversity values, including wetlands, savannas, grasslands, and shrublands. For instance, Brazilian savannas are well recognized as a biodiversity hotspots (Cerrado region) and it is the area that have lost most of their natural habitat in Brazil (Fig. 2), followed by the Pantanal (world’s largest tropical wetland). Note that wetlands provide multiple benefits, including biodiversity, carbon sequestration, and the conservation of the water cycle (and of the climate). Similarly, the Paraguayan Dry Chaco is quickly losing the native vegetation (Fig. 3).
Fig. 1. Examples of fast deforestation in two areas with high biodiversity value (in the Brazilian Amazon): Fishbone type deforestation pattern between 2000 and 2019 (top), and rectangular type deforestation pattern between 2013 and 2017 (bottom). The most deforested images (right) also show the smoke from fires associated with deforestation. Source: NASA Earth Observatory
Fig. 2. Native habitat lost in Brazil (as proportion of the area, between 1985 and 2020) by states (left) and by biogeographic regions (right). Source: elaborated from data in MapBiomas.org
Fig. 3. Deforestation in different South American ecosystems, from https://insights.trase.earth/insights/cop26-must-commit-to-zero-deforestation/
Fig. 4. Degradation of Indonesian ecosystems: Annual primary forest cover loss (in Mha) in Sumatra and Kalimantan (a), and remaining forest cover in 2001 and 2019 in peatland (b) and non-peatland (c) ecosystems. Deforestation of peatlands and artificial drainage are major cause of peatland fires in the area, and those fire are a major sources of CO2 emissions. From Nikonovas et al. (2020).
One of the major factors generating dynamic in Euro-Mediterranean landscapes is rural abandonment and the consequent expansion of oldfields [1]. This oldfields are been colonized by fast-growing flammable plants, and this has occurred now for few decades [2]. Thus wildfires often spread through oldfields [2], and in fact the increasing of oldfields is the main driver of the increased fire activity in Spain since the 70’s [2]. One example of Mediterranean crop is the Carob tree [1]. Carob trees (Ceratonia siliqua; Fabaceae) are evergreen tree that produce edible pods, used in the past mainly as animal fodder; currently it is also used as substitute for chocolate (pods are sweet; in fact they can be eaten raw from the tree).
A recent wildfire in Azuébar (August 2021, eastern Spain) burned a landscape that had many abandoned Carob tree groves. These trees burned at a range of seventies (degree of consumption). So perhaps, by looking at the carob trees across the burned landscape we could map the severity of a wildfire! In the past we estimated fire severity using the degree of consumption of the pine canopy (Pinus halepensis) [3].
Pictures taken in September 2021, one month after the Azuébar fire.
Figure 1. Carob tree unburned (top left) and burned with different severity: with leaves unconsumed (top right), with leaves consumed (mid left), with main trunk consumed (mid right), with most wood consumed (bottom, left), with everything consumed (bottom right). All pictures taken 1 month after a fire in Azuébar, eastern Spain. Photos: JG Pausas.
References
[1] Pausas JG, Bonet A, Maestre FT, Climent A. 2006. The role of the perch effect on the nucleation process in Mediterranean semi-arid oldfields. Acta Oecol. 29: 346-352. [pdf] [doi] [ScienceDirect]
[2] Pausas JG & Fernández-Muñoz S. 2012. Fire regime changes in the Western Mediterranean Basin: from fuel-limited to drought-driven fire regime. Climatic Change 110: 215-226. [doi | springer | pdf]
[3] Pausas, J.G., Ouadah, N., Ferran, A., Gimeno, T. & Vallejo, R. 2003. Fire severity and seedling establishment in Pinus halepensis woodlands, eastern Iberian Peninsula. Plant Ecol. 169: 205-213 . [doi | pdf]
The question of whether species are organised as collectives of integrated interacting assemblages (Clements’ community concept) or behave individualistically (Gleason’s community concept) is a century-old debate in ecology that is still unresolved. In a recent article, we are reconciling the two approaches [1].
The Gleasonian view suggests that communities are assembled by species that respond individualistically along environmental gradients and thus cannot form bounded units (Fig. 1A). However, in many world landscapes, for a given climate, strikingly different biomes with sharp boundaries co-occur forming landscape mosaics. These mosaics are typically formed by a closed biome (forests) and open (non-forest) biome (e.g., grassland, savanna, shrublands). These two alternative biome states (ABSs [2]) are maintained by different feedback processes and have radically different species with contrasting shade and disturbance tolerance traits [2].
Under the individualistic view of species along climatic gradients, the overlapping response curve along a climate gradient (Fig. 1A) may indicate plant coexistence (and potentially competitive interactions); however this is true only if they occur in the same biome (Fig. 1B). That is both Gleason’s individualistic view (within biome) and Clements’s organismic view (across biomes) are complementary; both perspective of community remain useful in ecology.
The consequence is that fitting species distribution models or using climate limits in modelling for projecting future species distributions are inappropriate for extensive regions with alternative biome states. One way to improve these predictions would be to consider the presence or absence of forest shade in the modelling [1].
Figure 1. Classical (Gleasonian) pattern of species response curves along a climate gradient (A), and the alternative pattern along the same climatic gradient when there are ABSs (B). Note that in the driest and the wettest section of the gradient, we find open (e.g., grassland) and closed (forest) biomes, respectively; but at intermediate levels of the gradient, both are possible depending mainly on the disturbance regimes and feedback processes [2). Thus, under the intermediate levels of the gradient, species that may seem to coexist when considering climate only (A) are not really coexisting but occurring in drastically different biomes (B). From [1].
References
[1] Pausas J.G. & Bond W.J. 2021. Alternative biome states challenge the modelling of species’ niche shifts under climate change. J. Ecol. 109: 3962-3971 [doi | pdf]
[2] Pausas J.G. & Bond W.J. 2020. Alternative biome states in terrestrial ecosystems. Trends Pl. Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
Este artículo apareció en TheConversation, ElDiario.es, LaMarea, y aquí, entre el 18 y 19 de agosto de 2021. Aquí incluyo una versión previa, ligeramente más larga que la publicada en el resto de medios.
En estos días de olas de calor se están produciendo grandes incendios en el Mediterráneo, especialmente en Turquía, Grecia, Italia, Argelia. Situaciones parecidas se están dando en otras partes mundo (California, Canada, Siberia, etc.). Y algo similar vivimos el verano pasado. Por ello se entiende que, repetidamente, me pregunten si todos estos incendios son consecuencia del cambio climático. La respuesta corta es que el cambio climático facilita los incendios (facilita la propagación del fuego y extiende la temporada de incendios), pero no determina que haya incendios. A continuación intento responder de manera más detallada.
Los ingredientes
Para que se den incendios se necesitan al menos tres ingredientes, que además deben darse de forma simultánea [1]. Estos ingredientes son: igniciones (naturales o humanas), vegetación densa y continua (combustible), y sequía. La relación de estos factores con los incendios no es lineal, sino de tipo umbral. Es decir, hay un nivel de igniciones, de continuidad de vegetación, y de sequía, a partir de los cuales la probabilidad de incendio aumenta de manera exponencial (se dispara).
Cuando se superan los tres umbrales se generan megaincendios de difícil control. Y estos umbrales varían con las condiciones meteorológicas. Concretamente, son muy bajos cuando las temperaturas son especialmente elevadas (olas de calor), la humedad baja, o los vientos son fuertes. Es decir, en estas condiciones, se necesita menos igniciones, menos combustible, y menos sequía para que se generen incendios. Por lo tanto, en esas condiciones particulares los incendios son mucho más probables, siempre y cuando haya igniciones y continuidad del combustible.
Efecto de las olas de calor y vientos fuertes (flecha roja) en modificar los umbrales que generan incendios. Fuente: [1]
El reciente incremento en sequías y olas de calor está asociado al cambio climático (ver informe IPCC 2021). Sin embargo, los incrementos en igniciones y en continuidad de la vegetación son bastante independiente del clima. El número de igniciones (tanto accidentales como provocadas) está muy relacionado con la actividad humana, y especialmente con actividades urbanas en zonas forestales o semiforestales. La continuidad de la vegetación está relacionada principalmente con el abandono rural y con plantaciones forestales densas sin una gestión apropiada.
El incremento de incendios en España en los años 70 y 80 se explica especialmente por el aumento en continuidad de la vegetación debido al abandono rural [2]. El cambio climático tuvo un papel poco relevante. A medida que dejamos que avance el cambio climático, el papel relativo del clima en los incendios aumenta. Hay que recordar que en España, y en muchos países europeos, la masa forestal está en aumento, a pesar de los incendios [3].
Por lo tanto, el incremento de las temperaturas, olas de calor y sequías facilita en gran manera los incendios, pero se requieren también igniciones y vegetación continua. Y eso es una buena noticia. Reducir las igniciones y generar discontinuidades en la vegetación es más sencillo que reducir el cambio climático (que también es necesario).
¿Qué podemos hacer?
La política de tolerancia cero a los incendios no ha funcionado en ningún país del mundo. Ni en países con presupuestos en extinción muy elevados. Eliminar los incendios de nuestros paisajes es imposible y contraproducente [4], especialmente en el marco del cambio climático. Debemos aceptar un cierto régimen de incendios y aprender a convivir con ellos.
El reto de la gestión es crear condiciones que generen regímenes de incendios sostenibles tanto ecológica como socialmente. Para conseguir esto no hay una receta sencilla ni única. Por ejemplo, no es lo mismo gestionar una zona donde los incendios se propagan por el paisaje principalmente gracias a vientos fuertes, que si lo hacen debido a la existencia de grandes extensiones forestales homogéneas [5]. En el primer caso, gestionar las igniciones puede ser lo más importante. En el segundo, la clave puede estar en gestionar el combustible.
Los incendios son especialmente peligrosos cuando se acercan a zonas semiurbanas (en la interfaz urbano-forestal) y es donde la gestión es más importante. Una manera de reducir los incendios es generar discontinuidades (horizontales y verticales) en la vegetación. Existen diversas herramientas para ello, tales como: realizar cortas y quemas prescritas, introducir herbívoros (salvajes o ganado), alternar sistemas forestales con cultivos y permitir que ardan los incendios que sean poco agresivos. Iniciativas como incentivar la actividad rural local (agricultura y ganadería extensiva) o la resilvestración (rewilding) pueden actuar en la buena dirección. Cada una de estas herramientas puede ser válida dependiendo del sitio y las condiciones, y dada la complejidad del sistema, puede ser importante utilizar una diversidad de herramientas. Ninguna de ellas elimina los incendios, pero reducen su probabilidad, su tamaño, y su intensidad.
En momentos de olas de calor o de vientos estivales fuertes (por ejemplo, durante los ponientes en Valencia) sería importante limitar las actividades humanas en el monte. Es decir, limitar el paso de vehículos y personas, incluyendo el acceso a segundas residencias situadas en entornos forestales. Si durante épocas de riesgo por pandemia se ha limitado la movilidad, quizá en momentos de máximo riesgo de incendios también se podría limitar la movilidad en zonas forestales y semiforestales. Esto es importante porque los incendios se producen cuando las igniciones coinciden con condiciones meteorológicas adversas en paisajes con suficiente vegetación. En esos escenarios, reducir las igniciones es clave.
También se podría limitar la interfaz urbano-forestal. Es decir, reducir la expansión de urbanizaciones y polígonos industriales en zonas rurales y naturales. Esta expansión, además de los efectos ambientales bien conocidos (en biodiversidad, especies invasoras, contaminación lumínica y visual, etc.), también constituyen una fuente de igniciones y hacen la vegetación más inflamable. Además, ponen en riesgo a personas e infraestructuras, y por lo tanto, convierten en catastróficos (socialmente) incluso a regímenes de incendios ecológicamente sostenibles. Los mecanismos para limitar estas zonas pueden ser diversos, incluyendo la recalificación de terrenos (a no urbanizables), o la implementación de tasas por construir en áreas con alto riesgo de incendios (pirotasas), entre otros. Y la planificación urbanística requiere considerar a los incendios, así como exigir estrategias de autoprotección alrededor de viviendas y la realización de planes de evacuación.
Otra medida importante es restaurar los humedales y otros ecosistemas litorales, ya que, a parte de los beneficios para la biodiversidad, mantienen el ciclo del agua y contribuyen a la conservación del clima [3]. La degradación de la costa (desecación de los humedales y la sobre-urbanización) contribuye a la reducción de la precipitación [3].
Y en cualquier caso, hay que reducir el consumo de combustibles fósiles. Esto ayudaría a frenar el aumento de CO₂ atmosférico, y así reducir la velocidad del cambio climático y la frecuencia de olas de calor. Y no solo por los incendios.
Referencias
[1] Pausas J.G. & Keeley J.E. 2021. Wildfires and global change. Frontiers in Ecology and Environment 19(7) [doi | wiley | pdf | brief for managers]
[2] Pausas J.G. & Fernández-Muñoz S. 2012. Fire regime changes in the Western Mediterranean Basin: from fuel-limited to drought-driven fire regime. Climatic Change 110: 215-226. [doi | springer | pdf]
[3] Pausas J.G. & Millán M.M. 2019. Greening and browning in a climate change hotspot: the Mediterranean Basin. BioScience 96:143-151. [doi | oup | blog | pdf]
[4] Pausas J.G. 2017. Acabar con los incendios es antinatural e insostenible. 20minutos (Ciencia para llevar), 13 Julio 2017. [20minutos | blog]
[5] Keeley, J.E. & Syphard, A.D. 2019. Twenty-first century California, USA, wildfires: Fuel-dominated vs. wind dominated fires. Fire Ecology, 15, 24.
Early humans and native cultures have used fire for clearing the ground from parasites and diseases, and some agricultural societies use fire to reduce livestock diseases [1,2]. In such cases, fire provide an ecosystem service to humans [2]. We recently asked to what extent this ‘cleaning effect‘ of fire is also observed in the wild, that is, whether wildfires may remove parasites and thus provide some benefits to wildlife [3]. To answer this question, we compared the presence of ecotoparasites (mites,Ophionyssus) in lizard populations of Psammodromus algirus living in recently burned areas with those in adjacent unburned areas, in eastern Spain. Our results suggest that many individuals of P. algirus survive fire (the smoke of the fire acts as a cue for quickly moving to safe microsites [4], e.g., crevices, under rocks, among roots; refugia [5]); and those that survived (and live in the postfire environment) have less ecotparasites (mites) than those living in unburned conditions. That is wildfires produce a ‘cleaning effect’ by reducing lizard ectoparasites in the postfire conditions. Fire, by disrupting the host-parasite interaction, provides a window of opportunity for lizards to avoid the negative effects of ectoparasites. We propose that wildfires likely fulfill a role in controlling vector-borne diseases and pathogens for other species, but this ecological effects have been largely overlooked.
Fig. 1. Probability of lizards infection by mites in relation to lizards’ size (snout-vent length) for recently burned areas (red) and for the corresponding paired unburned (blue). Lines are predicted values (and confidence intervals) from [3].
Fig. 2. Psammodromus algirus enjoying a clean (parasite-free) environment after a fire. Photo: Lola Álvarez-Ruiz
Fig. 3. Pictures of mites (genus Ophionyssus; in different stages) found under the scales of P. algirus, from [3]
Fig. 4. Art work on the paper by Josep Serra, 6/2021
References
[1] Pausas J.G. & Keeley J.E. 2009. A burning story: The role of fire in the history of life. BioScience 59: 593-601 [doi | OUP | pdf]
[2] Pausas J.G. & Keeley J.E. 2019. Wildfires as an ecosystem service. Front. Ecol. Environ. 17: 289-295. [doi | pdf]
[3] Álvarez-Ruiz L, Belliure J, Santos X., Pausas JG. 2021. Fire reduces parasite load in a Mediterranean lizard. Proceed. Royal Soc. B. [doi | pdf] New!
[4] Álvarez-Ruiz L, Belliure J, Pausas JG. 2021. Fire-driven behavioral response to smoke in a Mediterranean lizard. Behav. Ecol. [doi | oup | PDF]
[5] Pausas J.G. 2019. Generalized fire response strategies in plants and animals. Oikos 128: 147-153 [doi | pdf]
To generate wildfires we need some specific components (ignitions, fuel, and right conditions). Traditionally, this has been explained using the triangle approach [1] or the 4-switches approach [2]. We propose a more mechanistic model to explain wildfires, the threshold approach [3]. Under this view, wildfires occur when three thresholds are crossed (ignition, continuous fuel, and drought); and fire weather moves these thresholds to lower values and so it triggers the occurrence and spread of wildfires (Fig. 1). The size and duration of the fire largely depend on how long the fire weather lasts and the extent of the area containing suitable fuel.
Climate change increases the conditions conductive to fire, and thus it also increases the frequency in which some of these thresholds are crossed, extending the fire season and increasing the frequency of dry years. However, climate-related factors do not explain all the complexity of global fire regime changes as human factors are extremely important: humans shifts ignition patterns and modify fuel structure. Humans cause ignitions directly by accident or arson, but also indirectly by altering fuels that can make them more susceptible to ignitions (vegetation openings). Humans also modify fuel continuity, either reducing it (eg fragmentation) or increasing it (eg fire suppression). For instance, in many Mediterranean ecosystems, the drought threshold is crossed annually, and vegetation cover (fuel) is usually high enough for fire spread; thus, ignitions are a key factor. Larger populations of humans in the wildland-urban-interface will likely lead to increased ignition rates, and consequently higher probability of ignitions coinciding with extreme weather events to generate wildfires.
Fig. 1. Probability of fire occurrence vs ignitions; fire spread vs landscape fuel continuity; and, fuel flammability vs drought. In each of the three graphs, vertical lines indicate the thresholds. In all cases, fire weather (strong wind, high temperature, or low humidity) moves the curve (and the threshold) towards lower values (thick red arrow; i.e. , saturation is reached at lower values of the x axis), with the consequence of increasing the probability of an ignition resulting in a fire, the fire spread (for a given landscape configuration), and the flammability of the vegetation (fuel dries out quicker). The flow chart indicates the main factors affecting the fire drivers: growing population (in or near wildlands); fuel changes in the landscape (fragmentation, oldfields, fire exclusion, etc.); and climate change (driven by the increase in greenhouse gases). From [3].
Fig. 2. Top-left: Wind-driven fires in eastern Australia (8 Nov 2019). Top-right: Pyrocumulonimbus in Valencia (Spain; 15 Jul 2019). Bottom-left: Pyrocumulonimbus in La Pampa (Argentina; 29 Jan 2018). Bottom-right: hurricane Ophelia fueling fires in Portugal and Spain, and covering the UK with smoke (16 Oct 2017). Photos by NASA and ‘Servicio de Vigilancia Prevención incendios forestales Caperutxo, Alacant’. From: [3]
References
[1] Moritz et al. 2005. Wildfires, complexity, and highly optimized tolerance. P Natl Acad Sci USA 102: 17912–17.
[2] Bradstock RA. 2010. A biogeographic model of fire regimes in Australia: current and future implications. Global Ecol Biogeogr 19: 145–58.
[3] Pausas JG & Keeley JE 2021. Wildfires and global change. Front Ecol Environ. [doi | web | pdf]
Aquí una entrevista en la que explico es qué trabajamos, y claro, hablo de incendios forestales, biodiversidad, desertificación, la estrategia de biodiversidad de la Comisión Europea, etc.
Y a continuación otra entrevista realizada en motivo de la MedNight TV del 24 Septiembre 2021
On ANZAC Day (25 April; the national day of Australia, see wikipedia) I received the following query from an Australian colleague.
Apparently one of the few Australian soldiers that survived the Gallipoli battle (Turkey, 1915, WWI) picked up a pine cone (from a ‘Lone pine’), and took it back to Australia. The cone was kept on a shelf until 1933 when a horticulturalist extracted 5 seeds and germinated them. The seedlings were planted in botanic gardens; the 78-year-old trees now have special symbolic value to war veterans and the like (Fig. 1). The returned veterans are often referred to as ‘lone pine soldiers’ (and the battle, as the battle of the lone pine). My colleague asked me if the cone story had some credibility, for instance, would a pine seed germinate after 18 years?
Fig. 1. Plaque at the foot of pine tree in Kings Park, Perth, Western Australia. From [4]
The plaque in Kings Park (Fig. 1) suggest that the pine is a Pinus halepensis (Aleppo pine). However, the pines in Gallipoli peninsula (and in most Turkey) are of another species: Pinus brutia (Turkish Red Pine) [2]; the two species are very close related. So the cone that the soldier pick up should be a P. brutia, unless there was a plantation of P. halepensis there.
Pine seed, once there are out of the cone, do not last for long (mostly less than a year). However one of the characteristics of both P. halepensis and P. brutia is that they have some serotinous cones (the serotiny level is higher in P. halepensis than P. brutia, but both have a proportion of serotinous cones) [2,3]. Serotinous cones are those that remain closed after maturation, i.e., more than a year [1]. They accumulate for several years, forming a canopy seed bank; these cones open after the tree burns in a wildfire, and thus it is an adaptation to regenerate after fire [1]. At least in the case of P. halepensis, we have evidence that cones remaining close on the tree for many years; most cones open in less than 8 years, but some can last more than 15 years (Fig. 2). In addition we have evidence of cones remaining closed after harvest (at lest 12 years in my experience). This information is for P. halepensis, but could apply to P. brutia as well.
Fig. 2. Left: Frequency distribution of trees in relation to their maximum closed cone age for Pinus halepensis in eastern Spain. The gray pattern corresponds to the proportion of trees in population under high frequency of crown fires, the white to the proportion of trees where crown fires are rare. From [3]. Right: example of long-lived serotinous cones in P. halepensis in eastern Spain.
In conclusion, the Australian soldier may have picked up a serotinous cone, perhaps from a P. brutia. The seed were in the cone for 18 years and then extracted and planted. There are other accounts suggesting that several Australian soldiers took cones of both P. brutia, and P. halepensis (it’s difficult to understand why soldiers would collect pine cones after such a deadly battle, but this is another question …).
Currently most WWI memorials in Australia include a P. halepensis tree, a few a P. brutia. In the memorial cemetery near Gallipoli they planted (in the 1920s) a different pine, a Pinus pinea (stone pine, not native from Turkey, but from Italy and Spain). And in New Zealand (they shared with Australians the Gallipoli drama), the tree in ANZAC memorials includes Pinus radiata (from California) and Pinus canariensis (from Canary Islands, west Africa). That is, any of the ca. 120 pine species may do for a war memorial…
If Gallipoli has not been a fire-prone ecosystem, the pines would not be serotinous, the cones collected by the soldiers would not had kept the seeds, and we would not have pines in the Australian and New Zealand war memorials. So now, when you see a pine in a war memorial, just think about fire adaptations!
References
[1] Lamont BB, Pausas JG, He T, Witkowski, ETF, Hanley ME. 2020. Fire as a selective agent for both serotiny and nonserotiny over space and time. Crit. Rev. Plant Sci. 39:140-172. [doi | pdf | suppl.]
[3] Hernández-Serrano A., Verdú M., González-Martínez S.C., Pausas J.G. 2013. Fire structures pine serotiny at different scales. Amer. J. Bot. 100: 2349-2356. [doi | amjbot | pdf | supp.]
The evolutionary role of fire in animals has been poorly explored [1]. We recently conducted an experiment with the mediterranean lizard Psammodromus algirus to evaluate a possible adaptive response to fire [2]. Specifically we tested whether detecting a fire by smoke is adaptive for lizards living in fire-prone ecosystems. To do so we collected lizards from habitats with contrasted fire regimes (fire-prone shrublands vs. non-fire-prone dunes) and exposed them to smoke and to a false smoke (control) in a terrarium. We video-recorded the behavioral reaction to the treatments, and determined whether each individual detected the smoke and the intensity of the reaction. We found that in populations from fire-prone habitats, more lizards reacted to smoke, and their behavioral response was more intense than in lizard populations from non-fire-prone habitats. Our results suggest that the enhanced response to smoke may have been selected as adaptive behavior in fire-prone habitats to increase survival. Smoke has been considered a very important cue for enhancing germination in plants [3]; now we show that is also a cue for fire avoidance in animals. To our knowledge, this is the first evidence of smoke detection by reptiles in wild populations.
Lizards’ behavioral response (seconds of activity in one minute) to the treatments (Control or Smoke) and habitat (Fire-prone or Non Fire-prone). Predicted values (filled symbols with confidence intervals) and data (light colored symbols). Species: Psammodromus algirus. From [2].
References
[1] Pausas JG, Parr CL 2018. Towards an understanding of the evolutionary role of fire in animals. Evol. Ecol. 32: 113–125. [doi | pdf]
[3] Keeley & Pausas 2018. Evolution of ‘smoke’ induced seed germination in pyroendemic plants. S. Afr. J. Bot. 115: 251-255. [doi | pdf]; Moreira & Pausas 2018. Shedding light through the smoke on the germination of Mediterranean Basin flora. S. Afr. J. Bot. 115: 244-250. [doi | pdf]; Tormo et al. 2014. Field evidence of smoke-stimulated seedling emergence and establishment in Mediterranean Basin flora. J. Veget. Sci. 25: 771-777. [doi| pdf]; Moreira et al. 2010. Disentangling the role of heat and smoke as germination cues in Mediterranean Basin flora. Ann. Bot. 105: 627-635. [pdf | doi]
This video provides a powerful message from grasses to the world, written and narrated by William Bond: The Untold Story of Grasses.
Grasses created an open sunlit world, rich in plants and animals.
When you support a tree planting project … Stop. Think. Are the trees restoring a forest? Or are they destroying an ancient grassland? What might be lost?
Further readings
Bond WJ. 2019. Open Ecosystems: Ecology and Evolution Beyond the Forest Edge. Oxford University Press.
Pausas JG. & Bond WJ. 2019. Humboldt and the reinvention of nature. J. Ecol. 107: 1031-1037. [doi | jecol blog | pdf]
Pausas JG. & Bond WJ. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
Afforestation is not a solution to mitigate CO2 emissions [link]
In the Yunnan Province of China, P. yunnanensis occurs in two growth forms: as a tree (var. yunnanensis) and as a shrub (var. pygmea) [1]. The shrubby form occurs mainly in upper slopes and ridges (Fig. 1), where soils are poor and dry, and fires are likely. This shrubby pine is very interesting and quite unique among pines: it has serotinous cones (Fig. 2), and resprouts after fire from basal buds (Fig. 3), generating multi-stemmed shrubby pine populations [1]. Serotiny is common among pines [2] while resprouting is not [3], so pines with both serotiny and resprouting are rare; and having a multi-stemmed growth form is even rarer.
Fig. 1. Pine shrubland (Pinus yunnanensis var. pygmea) in Yunnan, China. Photo C. Luo [1]
Fig. 2. Pinus yunnanensis var. pygmea showing serotinous cones, Yunnan, China. Photo: W.-H. Su [1]
Fig. 3. Pinus yunnanensis var. pygmea resprouting from basal buds after a fire, Yunnan, China. Photo: JG Pausas [1]
References
[1] Pausas JG, Su W-H, Luo C, & Shen Z. 2021. A shrubby resprouting pine with serotinous cones endemic to Southwest China. Ecology [doi|pdf]
[2] He T, Pausas JG, Belcher CM, Schwilk DW, Lamont BB. 2012. Fire-adapted traits of Pinus arose in the fiery Cretaceous. New Phytol. 194: 751-759. [doi | wiley | pdf | suppl.]
[3] Pausas, J.G. 2015. Evolutionary fire ecology: lessons learned from pines. Trends Pl. Sci. 20: 318-324. [doi | sciencedirect | cell | pdf]
I’m compiling paintings about wildfires and burned landscapes. Here some examples (click the picture to enlarge it). More examples are welcome! (email), thanks!
George Catlin, 1832
Fred Williams, 1970
Cota Marqués, 2020
Philip Juras, 2003
Donald Ramsay
Josep Serra, 2012
The painters
George Catlin (1796–1872, USA) Fred Williams (1927–1982, Australia) Cota Marqués (Spain): painted during the lockdown 2020 Philip Juras (Southeast USA) Donald Ramsay (Australia) Josep Serra Tarragón (Spain)
Conservation of Earth’s biological diversity will be achieved only by recognition of the critical role of fire in shaping ecosystems.
Article:
Kelly LT, Giljohann KM, Duane A, Aquilué N, Archibald S, Batllori E, Bennett AF, Buckland ST, Canelles Q, Clarke MF, Fortin M-J, Hermoso V, Herrando S, Keane RE, Lake FK, McCarthy MA, Ordóñez AM, Parr CL, Pausas JG, Penman TD, Regos A, Rumpff L, Santos JL, Smith AL, Syphard AD, Tingley MW, Brotons L. 2020. Fire and biodiversity in the Anthropocene. Science 370 (6519): eabb0355. [doi | science | pdf | suppl.]
What is the difference between the top and bottom pine cones in this photo? This is a question I often ask to my new students in the first field trip; in this case, Beniardà fire, 2020 [link]
These cones are from Pinus halepensis and were collected after a wildfire in Beniardà (Alicante, E Spain; burned in Aug 2020).
Many of you reply correctly; here is the full answer:
Top cones: before the fire, they were open on on the tree, i.e., without seeds. Fire burn them, and so they are all black
Bottom cones (see also the picture below): before the fire they were closed (serotinous cones), and fire opened them facilitating seed dispersal. Note that they are unburned inside. These cones contribute to the postfire regeneration of the pine.
Serotinous cones in Pinus halepensis: before (left) and after a fire (right)
References
Lamont BB, Pausas JG, He T, Witkowski, ETF, Hanley ME. 2020. Fire as a selective agent for both serotiny and nonserotiny over space and time. Critical Reviews Plant Sci 39:140-172. [doi | pdf | suppl.]
Castellanos MC, González-Martínez S. & Pausas JG. 2015. Field heritability of a plant adaptation to fire in heterogeneous landscapes. Mol Ecol 24: 5633-5642. [doi | pdf | suppl.]
Hernández-Serrano A, Verdú M, González-Martínez SC, Pausas JG. 2013. Fire structures pine serotiny at different scales. Am J Bot 100: 2349-2356. [doi | amjbot | pdf | supp.]
Hernández-Serrano A, Verdú M, Santos-Del-Blanco L, Climent J, González-Martínez SC & Pausas JG. 2014. Heritability and quantitative genetic divergence of serotiny, a fire-persistence plant trait. Ann Bot 114: 571-577. [doi | pdf | suppl.]
Cork oak (Quercus suber) typically grows in relatively wet mediterranean environments [1]. However there are some cork oaks in arid climate; perhaps the population in the driest site is the small and isolated cork oak patch in Rambla de Talón (ca. 100 m asl, Ribera de Molina, Molina de Segura, Murcia, Spain; Fig. 1). It includes less than 100 individuals scattered in an area of sandy conglomerates (Fig. 3); the average rainfall is less than 300 mm. They are believed to have been planted in the past (when?), but their persistence in such arid conditions gives them a high added value. This population is much smaller and is located in a much drier conditions than the one in Pinet (Valencia) we mentioned some time ago [2,3].
Figure 1. Distribution of Cork oak (Quercus suber) in the Iberian Peninsula. Light grey is the species distribution; dark grey is the data from forest inventories; crosses are small isolated populations. In red is the population of Murcia. Map from [1].
Precipitation during the last spring was above average, and currently (end of October 2020) most oaks in Rambla de Talón look healthy and have some acorns. Of the 26 tree we look at, the number of acorns ranged from 0 (7 trees) to more than 400 acorns (2 trees), but most trees have less than 10 acorns (Fig. 2; median= 5 acorns). In addition, there is no evidence of recruitment from previous years. That is, if persistence of this population is desired, it would require some help for their regeneration. Given that they produce some acorns, restoration actions using local acorns is possible.
Fig. 2 Acorn production (October 2020) in 26 cork oak trees from Rambla de Talón, Ribera de Molina, Murcia.
Fig. 3. Views of the cork oak population in Rambla de Talón, Ribera de Molina, Murcia. Click the image to enlarge it.
References [1] Aronson J, Pereira JS, Pausas JG (eds). 2009. Cork Oak Woodlands on the Edge: conservation, adaptive management, and restoration. Island Press, Washington DC. 315 pp. [The book]
[2] Pausas JG, Ribeiro E, Dias SG, Pons J & Beseler C. 2006. Regeneration of a marginal Cork oak (Quercus suber) forest in the eastern Iberian Peninsula. J. Veget. Sci. 17: 729-738. [pdf | doi | wiley ]
Hummingbirds are among the most iconic birds of America, especially abundant in the tropics. They are very important pollinators (nectar feeders), thus their abundance and distribution are likely to affect hummingbird-pollinated plants, many of which are endemic or endangered. A recent review in western United States [1] suggests that most hummingbird species respond positively to wild or prescribed fire and thus, for the conservation of these emblematic birds, it is important to promote landscape mosaics, with early and mid postfire successional habitats, together with forest patches.
Here is a video by Contreras-Martínez et al. on hummingbirds and wildfires in Sierra de Manantlán Biosphere Reserve, Mexico [2]. The video is in Spanish; hummingbirds are ‘colibríes’ or ‘picaflores’
Grandes Guerreros: Colibríes y Fuego
Créditos del vídeo: Filmación y Edición: Carlos Armando Pacheco Contreras (Vidacinema) Productor: Sarahy Contreras Martínez Guión: Sarahy Contreras Martínez, Carlos Armando Pacheco Contreras & Oscar Cárdenas Hernández Narración: Cesar Híjar Tejada Investigación Científica: Sarahy Contreras Martínez (Colibríes) & Enrique Jardel Peláez (Fuego) Música Original: Erick Ríos Vázquez Mezcla de audio: Sognare Estudios Also available at: https://www.facebook.com/watch/?v=869506306897201
References [1] Alexander JD, Williams EJ, Gillespie CR, Contreras-Martínez S & Finch DM. 2020. Effects of restoration and fire on habitats and populations of western hummingbirds: A literature review. Gen. Tech. Rep. RMRS-GTR-408. Fort Collins, CO, USDA, https://www.fs.usda.gov/treesearch/pubs/59664
[2] Pausas JG 2016. Flammable Mexico. Internat. J. Wildland Fire 25: 711-713. [doi | pdf ]
Meteorologists call Cumulus (Cu) to cotton-like clouds, and Cumulonimbus (Cb) to denser and bigger clouds carried by powerful upward air currents. When these clouds are originated by a fire (or a volcano), we call them pyrocumulus (PyroCu) and pyrocumulonimbus (PyroCb) clouds.
Pyrocumuloninbus (= Cumulonimbus flammagenitus) are dense towering vertical clouds carried by powerful upward air currents generated by the heat of a wildfire [1]. These fires are also called plume-dominated fires, super-heated wildfires, or wildfire-driven thunderstorms. The origin is typically tied to very high and continuous fuel loads and extreme fire weather that produces great heat and strong convection currents. In most cases, they remain in the troposphere, but when the heat produced by the fire is very high, they can cross the tropopause and inject a large amount of smoke into the stratosphere; in those cases, wildfires contribute to the global carbon and nutrients redistribution [2]. These plumes often collapse when temperatures drop with altitude and create extreme winds. That is, these high intensity fires generate feedback processes between the atmosphere and the fire that produce strong surface winds, tornadoes, and even pyrogenic lightning ignitions that further expand the fire (firestorms).
Firestormsare wildfires with extreme, sometimes erratic behavior, typically beyond the capability of fire services to contain; the term has been used to describe very different fire types such as wind-driven fires (e.g., Santa Ana wind firestorms in California) and pyrocumulonimbus plume fires [1].
A
B
C
D
E
F
Examples of pyrocumulonimbus clouds (click in the picture for enlarge it).
A: Benaixama fire, Alicante (Spain), 7/2019, by Servicio de Vigilancia de Prevención de Incendios Forestales, Observatori Caperutxo, Generalitat Valenciana. B: Llutxent, Valencia (Spain), 8/2018, by Empar. C: Tasmania (Australia), 2013, by Janice James. D: Creek fire, California (US), 9/2020, by Thalia Dockery. E: La Pampa (Argentina), 1/2018, Earth Observatory, NASA. F: Funny river, Alaska (US), 5/2014, Earth Observatory, NASA.
References
[1] Pausas J.G. & Keeley J.E. 2021. Wildfires and global change. Front. Ecol. Environ. [doi | wiley | pdf]
[2] Pausas J.G. & Bond W.J. 2020. On the three major recycling pathways in terrestrial ecosystems. Trends Ecol. Evol. 35: 767-775. [doi | sciencedirect | pdf]
Despite the multiple evidence that afforestation is not a solution for mitigating the increased atmospheric CO2 [1], there are still lobbies and multimillionaire clubs willing to plant millions of trees at the global scale, and spreading myths about the benefits of trees and large afforestation programs. Recently (17 July 2020), William Bond gave a talk at Oxford University to bust these myths. Here is his talk, and below is a summary of the top 5 myths.
Myth 1. Forest are ancient, non-forests are caused by deforestation. There is evidence of ancient species-rich grasslands and shrublands in many parts of the world (from Cerrado in Brazil, to grasslands in Africa, shrublands in Mediterranea ecosystems, etc.). In fact animal grazers evolved long ago (long before humans could deforest) in grasslands. There are also evidences of many tropical forests that were thought to be ancient and are not (e.g., youtube). This myth has deep roots in the western culture [2].
Myth 2. Oxygen comes from trees: cutting down forest will deprive us of air to breath. Oxygen is more ancient than forests! The atmospheric concentration of oxygen during much of the evolutionary history of plants, before the rise of dense tropical forests, has been higher than current level (21%). Fire requires oxygen to burn, and there has been fire since early colonization of land plants [3]. Statements like the Amazon provides 20% of our oxygen are wrong; the Amazon consume about as much O2 as it produces; O2 is ancient, it doesn’t depend on trees (see details: link1 & link2). There are lots of reasons to preserve the Amazon, but running out of oxygen isn’t one of them.
Myth 3. Forests ‘make rain’: plant trees to get more water. W. Bond note that many city dwellers and some climatologists suggest that planting trees would increase water supply, but farmers, which have daily experience with land management, says that planting trees dries up rivers. A catchment experiment in South Africa unambiguously show that catchments with tree plantations get drier compared with those under natural shrublands (Wyk 1987). Maybe some catchments, given their size, climate and topography, may generate their own rainfall (as often suggested for the Amazon), but this doesn’t seems a general rule. Planting trees will not ‘make rain’, most likely will dry out the watershed (e.g., Wang et al. 2020).
Myth 4. The biggest store of terrestrial carbon is in tropical forests. Tropical forests store about 225 Pg C, while boreal soils store ca. 1300 Pg C. So, from the C perspective, it is more important to conserve boreal soils (peatlands, etc.) than tropical forests! Obviously tropical forest need to be conserved for their biodiversiy. But you better forget about planting trees, and start thinking in conserving boreal peatlands as their destruction would release high amount of CO2 to the atmosphere. See also: Friggens et al. 2020.
Myth 5. Forests equate with biodiversity. Many tropical forests are highly diverse, but there are examples where planting trees implies a loss of biodiversity (Abreu et al. 2017, Phifer et al. 2017). When comparing savannas and forest for the same rainfall, there are no differences in biodiversity (Murphy et al. 2016). In addition, many of the global biodiversity hotspots are open non-forest ecosystems or mosaics of forest and open ecosystems. So the myth cannot be hold. In fact, landscape mosaics of forest and non-forest are highly diverse landscapes [4].
[2] Pausas J.G. & Bond W.J. 2019. Humboldt and the reinvention of nature. J. Ecol. 107: 1031-1037. [doi | jecol blog | jgp blog | pdf]
[3] Pausas J.G. & Keeley J.E. 2009. A burning story: The role of fire in the history of life. BioScience 59: 593-601 [doi | OUP | pdf | post]
[4] Pausas J.G. & Bond W.J. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
— Update: a new paper that addresses this topic: Fleischman et al. 2020. Pitfalls of tree planting show why we need people-centered natural climate solutions. BioScience, doi: 10.1093/biosci/biaa094
Here is a new review paper on serotiny in plants [1]. Serotiny refers to prolonged storage of seeds in woody structures (cones or fruits) on the mother plant for several growing seasons. This implies an accumulation of a canopy seed bank with seeds from different. Serotiny confer fitness benefits in environments with frequent crown-fires, as the heat of the fires opens the woody structures and thus seeds are dispersed in the post-fire bed (where resource are abundant and competition is low). There are other ways by which some cone/fruit can be opened (e.g., dry warm winds), but the number of seeds released and the chance of successful recruitment are much lower than in postfire conditions, and thus fire provides higher fitness benefits for serotinous plants than any other cue factor. This is why most serotinous plants occurs in ecosystems with frequent crown fires. Serotiny has been studied mainly in pines [2-4] and Proteaceae, but we know at least 12 families and more than 50 genera with serotinous species [1]. And there is a diversity of serotinous structures (cones and fruits) in different genera and families (Fig. 1).
Figure 1. A) Callitris (Actinostrobus) pyramidalis (Cupressaceae); B) Postfire Cupressus sempervirens (Cupressaceae); C) Cone of Pinus patula; D) Postfire cones of Pinus radiata; E) Folicles of Hakea cyclocarpa (Proteaceae); F) Hakea stenocarpa follicle; G) Follicles of H. platysperma; H) Xylomelum angustifolium (Proteaceae) follicle; I) Lambertia echinata (Proteaceae) follicle; J) Cone of Banksia lemanniana (Proteaceae); K) B. hookeriana burning; L) Postfire B. leptophylla cone; M) Cluster of capsules of Leptospermum spinescens (Myrtaceae), inset: seeds of Callistemon (Melaleuca) teretifolius; N) Elongated cluster of sessile capsules of Callistemon citrinus (Myrtaceae), + seeds of Callistemon (Melaleuca) teretifolius; O) Woody capsules of Eucalyptus todtiana (Myrtaceae), Inset: capsules of Angophora hispida (Myrtaceae); P) Spikes of Connomois parviflora (Restionaceae); Q) Protea burchellii (Proteaceae); R) fruits of Protea burchellii; S) Cone of Allocasuarina torulosus (Casuarinaceae); T) Cone of Isopogon trilobus (Proteaceae); U) Cones of Petrophile brevifolia; V) Seedlings from serotinous H. polyanthema, B. attenuata and B. hookeriana in litter microsite. For more details see [1].
References
[1] Lamont BB, Pausas JG, He T, Witkowski, ETF, Hanley ME. 2020. Fire as a selective agent for both serotiny and nonserotiny over space and time. Critical Rev. Plant Sci. [doi | pdf]
[2] Hernández-Serrano A, Verdú M, González-Martínez SC, Pausas JG. 2013. Fire structures pine serotiny at different scales. Am. J. Bot. 100: 2349-2356. [doi | amjbot | pdf | supp.]
[3] Hernández-Serrano A, Verdú M, Santos-Del-Blanco L, Climent J, González-Martínez SC, Pausas JG. 2014. Heritability and quantitative genetic divergence of serotiny, a fire-persistence plant trait. Ann. Bot. 114: 571-577. [doi | pdf | suppl.]
[4] Castellanos MC, González-Martínez S, Pausas JG. 2015. Field heritability of a plant adaptation to fire in heterogeneous landscapes. Mol. Ecol. 24, 5633-5642. [doi | pdf | suppl.]
Plants are the largest biomass component of most terrestrial ecosystems, and litter decomposition is considered the dominant process by which nutrients return to plants. In a recent paper [1] we show that in terrestrial ecosystems, there are three major pathways by which plant biomass is degraded into forms that release nutrients again available to plants: microbial decomposition; vertebrate herbivory; and wildfires. These processes act at different spatial and temporal scales, have different niches, and generates different ecological and evolutionary feedbacks. The three processes can occur in a given ecosystem (competing for the same resource, biomass), but the relative importance of each varies with the micro- and macro-environmental conditions (see Figure below).
Wildfires and herbivory are two powerful biomass consumers; they generate feedback processes that maintain vegetation at states of lower biomass than would be expected from the physical environmental conditions (alternative vegetation states [2]). In addition, wildfires and herbivory also select for light-loving species with a set of adaptive traits to persist under these consumers [3,4]. That is, both herbivory and fire can influence the mix and attributes of plant species, while the mix and attributes of plants also influence the fire and grazing regimes. These ecological and evolutionary feedbacks make fire and herbivory distinct from other abiotic disturbances such as cyclones, landslides, avalanches, volcanoes, or floods, where plants may respond – but the disturbance will not change in response to these plant responses. That is, wildfires, herbivory, and microbial decomposition can be viewed as biotic processes that structure our ecosystems and the biosphere, at different temporal and spatial scales.
This holistic view in which microbes, herbivores, and wildfires play a joint role in the functioning of ecosystems contributes to a better understanding of the diversity of mechanisms regulating the biosphere.
Figure: Plant biomass and vegetation structure in terrestrial ecosystems are determined by three feedback processes: vertebrate herbivory (H), microbial decomposition (D), and wildfires (W). These three processes also interact with each other (e.g., competition for biomass; but positive interactions also exist). Relative importance of each of the three ecosystem pathways varies in the environmental space (niche), here defined by the water availability and soil fertility. Illustration by Dharmaberen Studio. From [1].
References
[1] Pausas J.G. & Bond W.J. 2020. On the three major recycling pathways in terrestrial ecosystems. Trends Ecol. & Evol. [doi | pdf]
[2] Pausas J.G. & Bond W.J. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
[3] Keeley J.E., Bond W.J., Bradstock R.A., Pausas J.G. & Rundel P.W. 2012. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management. Cambridge University Press. [the book]
[4] Bond, W. J. 2019. Open Ecosystems: Ecology and Evolution Beyond the Forest Edge. Oxford University Press.
The role of large herbivores in explaining broad-scale ecological pattern has probably been underestimated [1]. For instance, they are important in maintaining many landscapes and biomes across the world [2]. In a recent paper we show that the different history of megafauna abundance and extinctions in different continents has shaped the dominance of many plant traits [3]. Tropical Africa (paelotropics) and tropical South America (neotropics) are a clear example of contrasting megafauna history under similar environmental conditions. By comparing plant traits of woody species in different biomes (savannas and forests) and for the two different continents, we found that continent explain better the differences in plant traits than biome, climate, or soil, and that the differences between continents are consistent with the higher impact of large vertebrates in Africa than in South America. For instance, plants in African savannas tend to be more thorny and to have higher wood density, i.e., traits related to defense against megaherbivores. In contrast, South American savannas (Cerrado) harbor more species with underground bud banks (geoxyles [4]), and thicker protective barks, i.e., traits related to protect from wildfires [4,5].
Megafauna was certainly present in South America before the Holocene overkilling by humans, however, it is unlikely they live in the brazilian savannas (cerrado); their weak and vulnerable stems (low height growth, low wood density, and lack of spines), are unlikely to have evolved in regions with abundant browsers. We hypotheses that megafauna in South America was distributed in: 1) an open version of the current seasonally dry tropical forests (SDTF, e.g., Chaco) as the proportion of thorny species is similar to African savannas (and much higher than the cerrado); and 2) the subtropical grasslands, as they currently need to be maintained by humans due to the missing megaherbivores (landscape anachronism [1]).
Overall our results suggest that variation in plant traits in the tropics is unlikely to be fully understood without considering historical factors, especially the direct and indirect impacts of megafauna. Looking at plants and thinking on their megafauna history may provide novel insights for understanding vegetation patterns across the globe [1].
The effects of megafauna history (left: absence, right: present of megafauna) on plant functional traits of tropical woody species in different biomes (savannas and forests). Arrows between the dominant driver(s) (boxes) and traits (circles) indicate positive (blue) and negative (red) effects. From [3]
References
[1] Pausas JG & Bond WJ. 2019. Humboldt and the reinvention of nature. J. Ecol. 107: 1031-1037. [doi | jecol blog | jgp blog | pdf]
[2] Pausas JG & Bond WJ. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 25: 250-263. [doi | sciencedirect | cell | pdf]
[3] Dantas V & Pausas JG. 2020. Megafauna biogeography explains plant functional trait variability in the tropics. Glob. Ecol. & Biogeogr. [doi | pdf | data:dryad]
[4] Pausas JG, Lamont BB, Paula S, Appezzato-da-Glória B & Fidelis A. 2018. Unearthing belowground bud banks in fire-prone ecosystems. New Phytol. 217: 1435–1448. [doi | pdf | suppl. | BBB database]
[5] Pausas JG. 2017. Bark thickness and fire regime: another twist. New Phytol. 213: 13-15. [doi | wiley | pdf] & Pausas, J.G. 2015. Bark thickness and fire regime. Funct. Ecol. 29:317-327. [doi | pdf | suppl.]
On June 26, 2019 a wildfire started in Torre de l’Espanyol (Ribera de l’Ebre, Tarragona, Catalonia, Spain) and burned ca. 5000 ha. I recently visited the area (8.5 months postfire); the regeneration was still relatively low, as the winter has been quite dry, but many species were resprouting and some seedlings were starting to emerge. Here are some of the plants that were flowering: Platycapnos (Fumaria) cf. spicata, Muscari neglectum ssp. atlanticum, Fritillaria lusitanica (=F. boissieri, F. pyrenaica ssp. hispanica), Ophrys lupercalis (=O. forestieri; O. bilunulata was also flowering but not pictured below).
Platycapnos cf spicata
Muscari neglectum ssp atlanticum
Fritillaria lusitanica
Ophrys lupercalis
Photos by C. Guiote & JG Pausas; species identification by J. Oltra
En los últimos años hemos aprendido mucho sobre ecología y evolución de las plantas en relación a los incendios forestales [1]. Una de los puntos clave pare ello fue encontrar evidencias de que siembre, a lo largo de toda la historia evolutiva de las plantas, han habido incendios, incluyendo en las primeras comunidades vegetales que colonizaron el medio terrestre [2]. Y todo esto se sabe gracias a los avances en el estudio de los carbones fósiles. Uno de los estudiosos más importantes en paleo-fuegos es el geólogo Andrew Scott, quien ha dedicado gran parte de su vida a mirar carbones fósiles, y que en 2018 resumió sus investigaciones en un libro titulado Burning Planet: The story of fire through time [3]. Ahora tenemos la suerte de que ese libro se ha traducido al castellano, Planeta en llamas: la historia del fuego a través del tiempo (Galaxia Gutenberg, 2020). Creo que este libro es un buen complemento a mi libro sobre la ecología de los incendios forestales [4], y espero que ayude a la comunidad hispánica a entender mejor el papel ecológico, evolutivo, y ancestral que tiene el fuego en nuestros ecosistemas.
Referencias
[1] Ecology & Evolution in fire-prone ecosystems [enlace]
[2] Pausas J.G. & Keeley J.E. 2009. A burning story: The role of fire in the history of life. BioScience 59: 593-601 [doi | OUP | post | pdf]
[3] Scott A. 2018. Burning Planet: The story of fire through time. Oxford University Press. [versión española: Planeta en llamas: la historia del fuego a través del tiempo. Galaxia Gutenberg, 2020]
[4] Incendios forestales, una visión desde la ecología, CSIC-Catarata, 2012 [enlace]
Australia is a very flammable continent, and fires have been occurring there for millions of years. As a consequence, many plants and animals have developed adaptations and strategies to cope with recurrent fires. However, the current fire season in eastern Australia is really very severe, including not only very large fires but also high intensity firestorms. SE Australia has suffered other sever fire seasons in the past (an iconic example is the Black Friday bushfires in 1939). Why is this happening now? Here I’ve compiled key figures that help us to understand it.
In the last few years, Australia has been suffering an increase in temperature; on average, each year is hotter than the previous year (Fig. 1). In fact, 2019 was the warmest years, but also the driest year (with the lowest rainfall) ever recorded. December 2019, when most fires started, was climatically an extreme (Fig. 2). During the December heatwave (Fig. 3) some meteorological station (e.g., Penrith, near Sydney) recorded temperatures over 48oC, and the record of highest average maximum temperature for Australia was broken on two consecutive days (40.7 and 41.9oC on 17 and 18 Dec, respectively). January 4 was Canberra’s hottest day since records began (44oC). In such extreme weather conditions, ignitions easily become a wildfire (in fact, several of the wildfires started from a dry lightning), and fires spread very quickly in a vegetation that has been in a drought for many months. This generates not only huge areas burned (Fig. 4), but also very hot fires and strong uplift air columns that reach the stratosphere (pyrocumulonimbus). These are called firestorms. Firestorms produce there own winds and spread embers and the fire very fast; they even produce lightnings that generate additional wildfires. Firestorms produce extreme fire behaviour that is beyond the capacity of firefighters. In those fires, as it happens in volcanoes, the smoke reaches the stratosphere and circulates at very long distances (e.g., currently smoke from these fires has already reached South America).
The fire season has not ended yet. The ecological effects of these fires will depend on many factors (spatial variability of fire intensity, previous fires, species, etc…). The size and intensity of these fires suggest that they can have some negative consequences, but it is too early for any quantitative evaluation. Many plants are starting to resprout just few days after the fire, even under those drought conditions; some animals are leaving their hiding places, exploring the burned area, and carcasses are locally abundant suggesting patches of high animal mortality. We’ll see when will the rain come, and how plants and animals will respond. For humans, the consequences are catastrophic (fatalities, destruction of many infrastructures, smoke problems, etc.).
Fig. 1. In Australia, each year is hotter than the previous year, on average. From Australian Bureau of Meteorology
Fig. 2. December 2019 was climatically an extreme, unprecedented in relation to rainfall and temperature. Elaborated with data from Australian Bureau of Meteorology
Fig. 3. Global temperature in December 18th, 2019, as shown by Windy. Note also that part of the differences in temperature are due to the different time zones; i.e., middle of the day in Australia, night time in South America, and early morning in Africa.
Fig. 4. Major fires in south-east Australia by January 10th, 2020 (5,634,000 ha). From @eforestal [update Jan 18th: 6 millions ha]
Update (4/2020): For a map of the time-since-fire and fire severity across NSW fires, see: Bradstock et al. 2020, Global Change Biol., doi:10.1111/gcb.15111 (spoiler: most fires burned at relatively low severity!)
Update (2021): Pausas J.G. & Keeley J.E. 2021. Wildfires and global change. Frontiers Ecol. & Environ. 19: 387-395. [doi | wiley | pdf ]
There is growing interest in the application of alternative stable state (ASS) theory to explain major vegetation patterns in tropical ecosystems [1] and beyond [2]. In a recent paper [3] we introduced the theory as applied to the puzzle of non-forested (open) biomes growing in climates that are warm and wet enough to support forests (alternative biome states, ABSs; Fig. 1). Long thought to be the product of deforestation, diverse lines of evidence indicate that many open ecosystems are ancient. They have also been characterized as ‘early successional’ even where they persist for millennia. ABS is an alternative framework to that of climate determinism and succession (Table 1 below) for exploring forest/nonforest mosaics. Within climatic and edaphic constraints, consumers (fire and herbivores) can produce vastly different ecosystems from the climate potential and have done so for millions of years [4]. This framework explains not only tropical forest–savanna landscapes, but also other landscape mosaics across the globe (Fig. 2).
Fig. 1. Generalized feedback processes in fire-prone landscapes where open and closed biomes (e.g., a grassland and forest) are alternative stable states maintained by stabilizing feedbacks, while perturbations generate abrupt transitions among states (destabilizing factors). From: [3].
—
Fig. 2. Examples of multibiome landscape mosaics where closed forests alternate with open biomes (grasslands) that are maintained by mammal herbivory and fire. From: [3].
—
Table 1. Comparison of the three main dynamic processes assembling disturbance-prone communities and landscapes: classical (facilitation) succession, autosuccession, and ABS. From: [3].
References
[1] Dantas V.L., Hirota M., Oliveira R.S., Pausas J.G. 2016. Disturbance maintains alternative biome states. Ecol. Lett. 19: 12-19. [doi | wiley | pdf | suppl.]
[2] Pausas, J.G. 2015. Alternative fire-driven vegetation states. J. Veget. Sci. 26:4-6. [doi | pdf | suppl.]
[3] Pausas J.G. & Bond W.J. 2020. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. [doi | sciencedirect| pdf]
[4] Pausas J.G. & Bond W.J. 2019. Humboldt and the reinvention of nature. J. Ecol. 107: 1031-1037. [doi | jecol blog | jgp blog | pdf]
España es un país muy diverso, con una gran variedad de climas, paisajes, comidas, bailes, lenguas, hablas, etc.; una diversidad generada por su peculiar situación geográfica, su heterogeneidad topográfica y la variedad de acontecimientos ocurridos a lo largo de su historia. Esto ha conferido a la población una elevada diversidad cultural, de ideas y puntos de vista. Sólo un gobierno plural y diverso, capaz de aceptar las diferencias, podrá gestionar correctamente y preservar la diversidad de este país. El bipartidismo difícilmente puede representar la diversidad española.
En la figura (abajo) vemos la evolución de la diversidad política en España, calculado a partir la distribución de los escaños de los diferentes partidos (en azul), para todas las Elecciones Generales al Congreso realizadas durante la democracia. El 15M (mayo 2011) sirvió para revitalizar la diversidad política del Congreso, tras muchos años de decaimiento (ver figura, en azul).
Tras las últimas elecciones generales en noviembre de 2019, en España tenemos un Congreso con la mayor diversidad de ideas políticas de la historia de la democracia (ver figura, linea azul); y esperamos que sea capaz de gestionar bien la diversidad del país. La diversidad del Congreso es prácticamente la misma que después de las elecciones de abril de 2019 (como era esperable), y sin embargo, ese Congreso fue incapaz de gestionar esa diversidad, seguramente por la falta de experiencia democrática, es decir, falta de capacidad de diálogo, de capacidad para aceptar las diferentes opiniones, y de excesivo deseo de poder de los lideres. Esperemos que el nuevo Congreso, y el nuevo gobierno que ahora empieza, hayan aprendido la lección.
Cabe señalar que la diversidad de opciones políticas de los votantes españoles, calculadas a partir de los votos y las abstenciones (linea roja de la figura) es más elevada que la diversidad política que se refleja en el Congreso (escaños; linea azul). El sistema electoral español hace que se pierda un parte nada desdeñable de la diversidad política de los ciudadanos. Con la llegada del 15M se ha visto reducida esa pérdida de diversidad (la linea azul se acerca más a la roja). Aproximadamente el 30% de los censados decidieron no contribuir a confeccionar la composición del Congreso (abstenciones, votos en blanco, votos nulos), y por lo tanto, contribuyen a esa pérdida de diversidad en el Congreso. La figura también refleja que pequeños cambios de opinión en los votantes a lo largo del tiempo (linea roja) se reflejan de manera amplificada en el Congreso (linea azul).
Figura: Valores de diversidad política (expresada con el índice de diversidad de Shannon-Weaver) calculado a partir de las opciones de los votantes (incluye votos a partidos, blancos, nulos y abstención; en rojo) y calculado según la distribución de los escaños resultantes (azul), para cada una de las Elecciones Generales al Congreso (España) realizadas durante la democracia (elaborado a partir de la base histórica de resultados electorales del Ministerio del Interior).
Nota aclaratoria: en las entradas previas sobre diversidad política en este blog, la diversidad política de los votantes (linea en rojo) se calculaba en base a la distribución de los votos a los distintos partidos. En esta entrada, y par reflejar un poco mejor la diversidad de las personas con derecho a voto, se ha calculado no sólo en base a los votos a los partidos, sino también incluyendo los votos en blanco, los nulos y las abstenciones.
I’m compiling paintings about wildfires and burned landscapes. Here some examples (click the picture to enlarge it). I would appreciate additional examples (email), thanks!
George Catlin, 1832, Missouri
Eugene von Guerard, 1857
Vasily Polenov, 1881
Fred Williams, 2004, Australia
Tom Thomson, 1915
Charles Burchfield, 1920
Rick Amor, 2014
Josep Serra Tarragón
Donald Ramsay, 2013, Australia
The painters
George Catlin (1796–1872, USA) Eugene von Guerard (1811–1901, Austria, Australia) Vasily Polenov (1844–1927, Russia) Tom J. Thomson (1877–1917, Canada) Charles E. Burchfield (1893–1967, USA) Fred Williams (1927–1982, Australia) Rick Amor (1948-, Australia) Donald Ramsay (Australia) Josep Serra Tarragón (Spain)
Ecosystems in southern Chile are not considered among the typical fire-prone ecosystems such as tropical savannas or mediterranean ecosystems. However, natural wildfires do occur (and has occurred since long ago), during drought periods, and are part of the ecological processes of the region. Here are some examples I have just visited.
Fitzroya cupressoides (alerce in Spanish, lahuán or lawal in Mapuche) is a shade-intolerant long-lived conifer native to the Andes of southern Chile and Argentina. Fitzroya is a monotypic genus in the cypress family. It often coexist with shade-tolerant species of Nothofagus (e.g., N. nitida). The bark of Fitzroya is relatively thick, and postfire tree survival depends on the intensity of fire; fire intensity in these ecosystems is typically patchy and some trees, especially large trees, do survive (Fig. 1 below and [1]). In fact, wildfires remove the shade-tolerant trees and open the space for Fitzroya which regenerates vegetatively (from root suckers) or from seeds coming from the surviving trees. Without wildfires, it would be hard for Fitzroya to compete with shade-tolerant broad-leaved trees.
Fig. 1. Dead and surviving Fitzroya cupressoides trees after fire in Parque Nacional Alerce Costero, Chile
Araucaria araucana (araucaria) is a conifer, considered a living fossil, native to central and southern Chile and western Argentina. It is a non-flammable tree (sensu [2]) because it typically self-prune their lower branches, the crown is quite open, it has a thick bark, and their foliage is hard and difficult to burn. This very low flammability allows Araucaria to survive even in flammable environments [2]. For instance, it occurs in shrublands of Nothofagus antartica (ñirre; see Fig. 2 below); this Nothofagus is a flammable multi-stemmed shrub that has a strong basal resprouting ability. This shrubland burn with some frequency but most Araucaria tree do not get burnt (fire can leave some scars in the trunk, see Fig. 3 below and dendroecological analysis in [3]). Araucaria araucana also growth in dens forests either as dominant tree or with other trees such as Nothofagus pumilo (lenga); such forest rarely burn and the regeneration of araucaria is based on gap dynamics. In fact, the two ecosystems (the shrublands of N. antartica, and the forests of N. pumilo) are an example of alternative biome states [4,5].
Fig. 2. Araucaria araucana growing in a shurbland of Nothofagus antartica (ñirre) in the foothills of the Lanín volcano, Chile
Fig. 3. Fire scars in three araucaria alive trees in the foothills of the Lanín volcano, Chile
References
[1] Lara A, Fraver S, Aravena JC & Wolodarsky-Franke A. 1999. Fire and the dynamics of Fitzroya cupressoides (alerce) forests of Chile’s Cordillera Pelada. Ecoscience, 6, 100-109.
[2] Pausas JG, Keeley JE, Schwilk DW. 2017. Flammability as an ecological and evolutionary driver. J. Ecol. 105: 289-297. [doi | wiley | pdf] [post-1 | post-2]
[3] González ME, Veblen TT & Sibold JS. 2005. Fire history of Araucaria–Nothofagus forests in Villarrica National Park, Chile. J. Biogeogr. 32:1187-1202.
[4] Pausas JG. 2015. Alternative fire-driven vegetation states. J. Veget. Sci. 26:4-6. [doi | pdf | suppl.]
[5] Pausas JG & Bond WJ. 2020. Alternative biome states in terrestrial ecosystems. Trend Plant Sci. [postprint]




































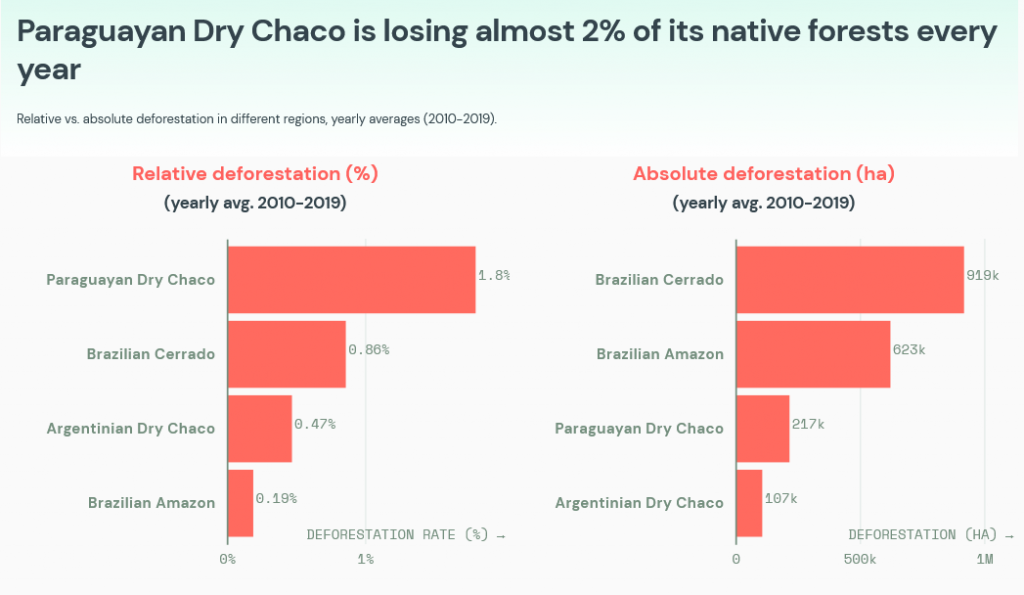





































































{kind=link}